Olá pessoal, hoje eu trago um tema introdutório em microbiologia: a morfologia e estrutura de bactérias.
A ciência da microbiologia é inteiramente destinada ao estudo dos microrganismos e do modo como eles funcionam, especialmente as bactérias, um grupo extenso de células muito pequenas que possuem grande importância básica e prática. Entretanto, a microbiologia também se debruça sobre os vírus e os fungos.
Os microrganismos podem ser tanto benéficos quanto prejudiciais para os seres humanos, embora a quantidade de microrganismos benéficos (ou até mesmo essenciais) seja bem maior do que os prejudiciais. Agricultura, alimentação, energia e meio ambiente são todos impactados de diversas maneiras pelos microrganismos.
Microrganismos vivem em populações que interagem com outras populações formando comunidades microbianas. As atividades dos microrganismos em comunidades microbianas podem afetar consideravelmente as propriedades químicas e físicas de seus hábitats. A biomassa microbiana na Terra excede aquela dos organismos superiores, embora a maioria das células microbianas surpreendentemente residir na subsuperfície profunda terrestre e oceânica.
A célula procariota e a célula eucariota
A análise da estrutura interna da célula revela dois padrões, chamados de procariota e eucariota.
Os procariotas incluem os domínios Bacteria e Archaea e consistem em células pequenas e estruturalmente bastante simples.
Os eucariotas são caracteristicamente maiores que os procariota e contêm uma variedade de estruturas citoplasmáticas envoltas em membranas chamadas de organelas. Essas incluem, mais proeminentemente, o núcleo que contém o DNA, mas também mitocôndrias e cloroplastos, organelas especializadas no fornecimento de energia para a célula, além de diversas outras organelas.
Os microrganismos eucariotas incluem algas, protozoários e outros protistas, bem como os fungos. As células de plantas e animais também são eucarióticas. Apesar das diferenças estruturais claras entre procariotos e eucariotos, a palavra “procariota” não implica um parentesco evolutivo. Como será visto na próxima seção, embora espécies de bactérias e arqueias possam parecer similares, elas não estão intimamente relacionadas do ponto de vista evolutivo.
Os vírus foram propositalmente excluídos desta discussão porque, embora eles se assemelhem a células de diversas maneiras, vírus não são células e representam uma categoria especial de microrganismos. São ditos parasitas intracelulares obrigatórios, pois por serem acelulares não possuem metabolismo próprio e necessitam obrigatoriamente parasitar uma célula para fazer uso de seu metabolismo e gerar novas unidades virais.
Breve histórico da vida na Terra e os três domínios da vida:
A vida celular encontrava-se presente na Terra há cerca de 3,8 bilhões de anos (bia). As cianobactérias iniciaram a lenta oxigenação da Terra há cerca de 3 bia, porém os atuais níveis de O₂ na atmosfera não foram alcançados antes dos últimos 500 a 800 milhões de anos. Os eucariotos são células nucleadas e incluem organismos microbianos e multicelulares.
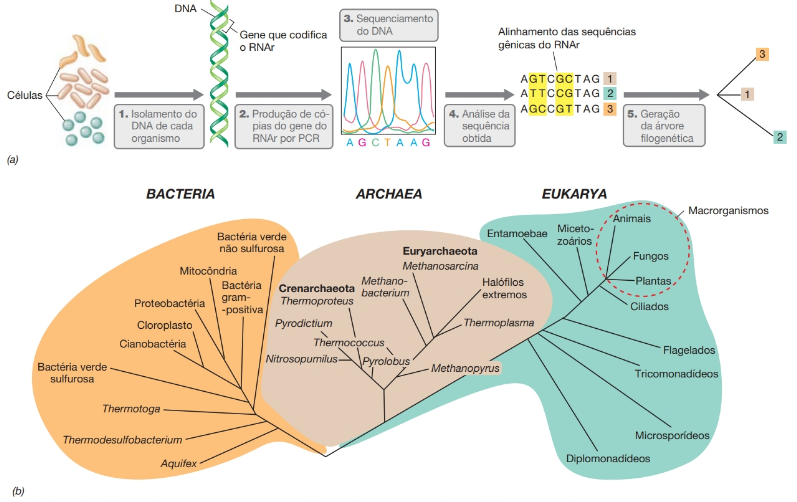
Propostos por Carl Woese, os três domínios dos organismos celulares são: Bacteria, Archaea e Eukarya. Archaea e Eukarya divergiram muito antes de as células nucleadas com organelas (em parte “eucariotos modernos”) aparecerem no registro fóssil. LUCA (do inglês, last universal common ancestor, ou último ancestral universal comum). Essas relações filogenéticas entre os 3 domínios foram descobertas em função da filogenia do gene do RNA ribossomal.
Relações evolutivas e a árvore da vida filogenética:
A tecnologia por trás da filogenia do gene do RNA ribossomal:
- 1. O DNA é extraído das células;
- 2. Cópias do gene que codifica o RNAr são produzidas por meio da reação em cadeia da polimerase (PCR);
- 3,4. O gene é sequenciado e a sequência obtida é alinhada com sequências de outros organismos;
- 5. Um algoritmo de computador realiza comparações de pares em cada base e gera uma árvore filogenética, que retrata as relações evolutivas.
No exemplo mostrado, as diferenças na sequência são destacadas em amarelo, sendo as seguintes: organismo 1 versus organismo 2, três diferenças; 1 versus 3, duas diferenças; 2 versus 3, quatro diferenças. Além disso, os organismos 1 e 3 possuem parentesco mais próximo do que 2 e 3 ou 1 e 2. (b) A árvore filogenética da vida. A árvore mostra os três domínios dos organismos e alguns grupos representativos em cada domínio.
Bactérias:
• Bactérias existem há mais do que 3,5 bilhões de anos;
• São unicelulares e procarióticas;
• São microscópicas, geralmente < 8μm;
• Só as colônias são vistas a olho nu ou bactérias gigantes;
• Devido a sua estrutura simples, as bactérias podem sobreviver em todos ambientes da terra;
• Podem ser encontradas no ar, solo, água, vulcão, mar profundo, fontes quentes, gelo, sal, pele dos homens etc.;
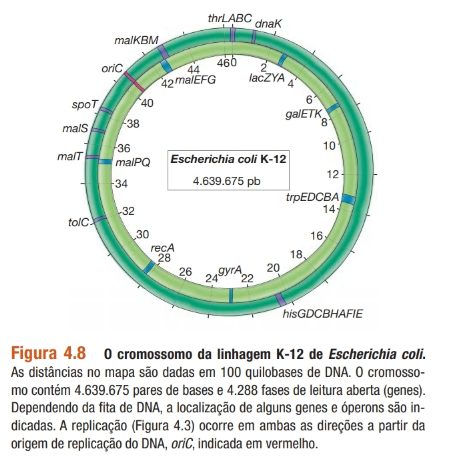
Cromossomo, nucleoide e plasmídeos:
Plasmídio: moléculas circulares de DNA capazes de se reproduzir independentemente do DNA cromossômico. DNAs menores. Mais encontrados em Gram-negativas. Não são essenciais, mas quando
presentes, conferem vantagem seletiva.(Ex.: resistência aos antibióticos);
Nucleoide: região que concentra o material genético;
Cromossomo: 1 único cromossomo, circular.
Os processos vitais das células são controlados pelo seu conjunto de genes, o genoma. Um gene é um segmento de DNA que codifica proteínas ou moléculas de RNA. O genoma é o projeto de vida de um organismo; as características, atividades, e sobrevivência de uma célula são controladas pelo seu genoma. Os genomas de procariotos e eucariotos são organizados diferentemente.
Em eucariotas, o DNA está presente na forma molecular linear, dentro de uma membrana fechada, chamada de núcleo.
Em contrapartida, o genoma de bactérias e arqueias está presente na forma de um cromossomo circular fechado (alguns procariotas apresentam cromossomo linear). O cromossomo se agrega dentro da célula para formar o nucleoide, uma massa visível ao microscópio eletrônico.
A maioria dos procariotas possui apenas um cromossomo, mas muitos podem apresentar, também um ou mais pequenos círculos de DNA diferentes do DNA cromossômico, chamados de plasmídeos. Os plasmídeos contêm genes que conferem determinada propriedade especial à célula (como um metabolismo diferenciado ou resistência a antibiótico) em vez de genes essenciais indispensáveis em todas as condições de crescimento. Essas características contrastam com os genes cromossômicos, a maioria dos quais são necessários para a sobrevivência básica.


Morfologia celular bacteriana:
Ao lado de cada desenho das células existe uma fotomicrografia de contraste de fase demonstrando a morfologia. Coco (diâmetro celular na fotomicrografia, 1,5 mm); bacilo (1 mm); espirilo (1 mm); espiroqueta, (0,25 mm); bactérias com brotamento (1,2 mm); bactérias filamentosas (0,8 mm). Todas as fotomicrografias são de espécies de bactérias. Nem todas estas morfologias são conhecidas entre as arqueias.
Endósporo:
• Em condições desfavoráveis ao seu crescimento algumas bactérias formam esporos, que podem sobreviver milhões de anos.
• Um esporo resulta da perda de água da célula e da formação de uma parede grossa e resistente em todo o citoplasma desidratado;
• O esporo consegue suspender completamente a sua atividade metabólica, sobrevivendo a situações adversas como calor intenso e falta de água;
• Os esporos bacterianos são muito importantes para medicina e indústria alimentícia, pois são resistentes ao calor e à esterilização química, podendo contaminar alimentos e provocar doenças - como antraz (Bacillus anthracis), tétano (Clostridium tetani) e botulismo (Clostridium botulinum);
• Uma das formas de eliminar definitivamente os esporos é a esterilização em autoclave, que consiste no uso de temperaturas de 120°C.
Determinadas espécies de Bacteria produzem estruturas denominadas endósporos, durante um processo denominado esporulação. Os endósporos (o prefixo “endo” significa “no interior”) são células altamente diferenciadas que exibem extrema resistência ao calor, produtos químicos fortes e radiação.
Os endósporos atuam como estruturas de sobrevivência e permitem ao organismo resistir a condições de crescimento adversas, incluindo, mas não limitadas, a extremos de temperatura, dessecamento ou carência nutricional. Desse modo, os endósporos podem ser considerados como o estágio latente de um ciclo de vida bacteriano: célula vegetativa -> endósporo -> célula vegetativa. Os endósporos ativados são estimulados a germinar quando colocados na presença de nutrientes específicos, como determinados aminoácidos.
Os endósporos também são facilmente dispersos pela ação do vento, da água ou por meio do trato gastrintestinal de animais. As bactérias formadoras de endósporo são encontradas predominantemente no solo, sendo as espécies de Bacillus os representantes mais bem estudados.
 |
| Fotomicrografias de contraste de fase ilustrando tipos morfológicos e localizações intracelulares dos endósporos, em diferentes espécies de bactérias formadoras de endósporos. Os endósporos aparecem brilhantes pela microscopia de contraste de fase. |
Durante a formação do endósporo, uma célula vegetativa é convertida a uma estrutura resistente ao calor, refringente à luz, e que não exibe crescimento (Figura 2.43). As células não esporulam quando se encontram em crescimento ativo, mas somente quando o crescimento cessa devido à exaustão de um nutriente essencial. Assim, células de Bacillus, uma típica bactéria formadora de endósporo, interrompem o crescimento vegetativo, iniciando o processo de esporulação quando, por exemplo, um nutriente essencial, como carbono ou nitrogênio, torna-se limitado.
Um endósporo pode permanecer dormente durante anos, porém pode converter-se novamente em uma célula vegetativa de forma relativamente rápida. Esse processo envolve três etapas: ativação, germinação e extrusão (Figura 2.44). A ativação acontece quando os endósporos são aquecidos por alguns minutos, a uma temperatura elevada, porém subletal. Em seguida, os endósporos ativados são estimulados a germinar quando colocados na presença de nutrientes específicos, como determinados aminoácidos. A germinação, um processo geralmente rápido (da ordem de alguns minutos), envolve a perda da refringência microscópica do endósporo, maior capacidade de coloração por corantes, e perda da resistência ao calor e produtos químicos. O estágio final, a extrusão, envolve um intumescimento visível decorrente da captação de água, e síntese de RNA, proteínas e DNA. A célula vegetativa emerge a partir do endósporo rompido iniciando seu crescimento, mantendo-se em crescimento vegetativo até que os sinais ambientais novamente desencadeiem a esporulação.



 |
Estágios da formação de um endósporo. Os estágios são definidos com base em estudos genéticos e análises microscópicas da esporulação em Bacillus subtilis, o organismo-modelo para estudos de esporulação.
|
Cápsula:
• Camada de consistência viscosa formada por polissacarídeos que reveste a parede celular em algumas bactérias;
• Goma xantana é um polissacarídeo extracelular produzido pelas bactérias do gênero Xanthomonas, sobretudo Xanthomonas campestris;
• Relaciona-se com a capacidade de aderência;
• É encontrada principalmente nas bactérias patogênicas.
Tradicionalmente, se a camada é organizada em uma matriz compacta, que exclui partículas pequenas, como a tinta nanquim, é denominada cápsula. Essa estrutura é facilmente visível por microscopia óptica se as células forem tratadas com tinta nanquim e também podem ser vistas ao microscópio eletrônico. Em contrapartida, se a camada é mais facilmente deformável, ela não excluirá as partículas e será de visualização mais difícil; essa forma é denominada camada limosa. As cápsulas normalmente aderem firmemente à parede celular, e algumas são até mesmo ligadas covalentemente ao peptideoglicano. As camadas limosas, contrariamente, ligam-se frouxamente, podendo ser perdidas da superfície celular.
Camadas superficiais externas têm várias funções. Os polissacarídeos de superfície auxiliam na ligação dos microrganismos às superfícies sólidas. Como será visto posteriormente, os microrganismos patogênicos, que penetram no corpo por vias específicas, geralmente realizam esse processo ligando-se inicialmente a componentes de superfície específicos dos tecidos do hospedeiro; essa ligação frequentemente é mediada por polissacarídeos de superfície da célula bacteriana. Quando surge a oportunidade, bactérias de todos os tipos normalmente se ligam a superfícies sólidas, algumas vezes formando uma espessa camada de células, denominada biofilme. Os polissacarídeos extracelulares também desempenham um papel essencial no desenvolvimento e na manutenção dos biofilmes.
Além da ligação, as camadas de superfície externas também podem desempenhar outras funções. Estas incluem a atuação como fatores de virulência em certas doenças bacterianas, e a prevenção da desidratação das células. Por exemplo, os agentes causadores das doenças antraz e pneumonia bacteriana – Bacillus anthracis e Streptococcus pneumoniae, respectivamente – apresentam, cada um, uma cápsula espessa de proteína (B. anthracis) ou polissacarídeo (S. pneumoniae). As células encapsuladas dessas bactérias evitam a sua eliminação pelo sistema imune do hospedeiro devido ao fato de que as células imunes, que, caso contrário, reconheceriam esses patógenos como invasores e provocariam a sua destruição, são impedidas de fazê-lo devido à presença da cápsula bacteriana. Adicionalmente a esse papel na doença, as camadas de superfície externas, praticamente de qualquer tipo, são capazes de se ligar a água e são passíveis de proteger a célula da dessecação em períodos de seca.

A goma xantânica é feita com substancias poliméricas extracelulares da bactéria Xanthomonas campestris.
Membrana plasmática:
• Camada que delimita o espaço da célula ,mantém condições adequadas para que ocorram as reações metabólicas, seleciona o que entra e sai da célula, ajuda a manter o formato celular;
• Diferencia-se da dos seres eucariontes por não conter esteróis, sendo uma estrutura fluida, que permite a mobilidade de proteínas (permeases, enzimas respiratórias, enzimas hidrolíticas etc.);
• Proteínas (60%) imersas numa bicamada fosfolipídica (40%);
• Transporte de solutos: difusão facilitada, transporte ativo e translocação de grupo;
• Produção de energia: transporte de elétrons e fosforilação oxidativa;
• Biossíntese de Macromoléculas: lipídios de membrana, peptidoglicano, ácido teicóico, ácido lipoteicóico, polissacarídeos extracelulares (LPS) e duplicação do DNA;
• Excreção de enzimas hidrolíticas;
• Taxia: quimiotaxia e fototaxia.
A membrana citoplasmática circunda o citoplasma e o separa do ambiente externo. Se a membrana citoplasmática estiver comprometida, a integridade celular será destruída, havendo extravasamento do citoplasma para o ambiente, provocando a morte da célula. A membrana citoplasmática é estruturalmente fraca e confere pouca proteção contra a lise osmótica, contudo, é a estrutura ideal para executar a sua principal função na célula: permeabilidade seletiva.
A membrana citoplasmática é uma barreira de permeabilidade altamente seletiva, constituída de lipídeos e proteínas que formam uma bicamada, internamente hidrofóbica e externamente hidrofílica. Em contrapartida a bactérias e eucariotos, em que os ácidos graxos são ligados ao glicerol por uma ligação éster, arqueias contêm lipídeos unidos por ligações éter e algumas formam uma membrana em monocamada, em vez de em bicamada.
A principal função da membrana citoplasmática é a permeabilidade, transporte e conservação de energia. Para acumular nutrientes contra um gradiente de concentração, sistemas de transporte são utilizados e caracterizados por sua especificidade e efeito de saturação.
Pelo menos três sistemas de transporte são bem caracterizados em procariotos. O transporte simples consiste somente em uma proteína transportadora transmembrânica, a translocação de grupo envolve uma série de proteínas no evento de transporte, e o sistema de transporte ABC consiste em três componentes: uma proteína de ligação ao substrato, um transportador integrado à membrana e uma proteína que hidrolisa ATP. Todos esses sistemas de transporte conduzem o evento em si utilizando a energia da força próton-motiva, do ATP, ou de algum outro composto orgânico rico em energia.
Parede celular:
• Envoltório extracelular rígido responsável pela forma da bactéria, com a função de proteger a célula contra agressões físicas do ambiente. Não possui celulose como as das células vegetais.
• Funções: manutenção da forma celular, suportar a elevada pressão osmótica
e auxiliar na divisão celular;
• Estrutura: Gram-positivas: 15-50% de mureína; Gram-negativas: aprox. 5% de mureína;
• Peptideoglicanos: peptídeos + heteropolissacarídeos; componente de paredes celulares bacterianas; heteropolímero de unidades alternantes de N-acetilglucosamina e ácido N-acetilmurâmico (β1->4).
Bactérias Gram-positivas:
Parede composta de até 90% peptoglicano, pode chegar a 50% do peso da bactéria. Contém ácidos teicóicos e ácidos lipoteicóicos.
Propriedades:
- Facilitar a entrada e saída de cátions;
- Regular atividade de autolisinas;
- Sítios receptores de bacteriófagos;
- Adesinas ao epitélio do hospedeiro;
- São antígenos celulares – permitem a identificação sorológica;
- Foi descoberto através da criomicroscopia eletrônica, que um espaço periplasmático muito menor está presente na bactéria Gram-positiva;
- Coram-se de roxo pela Coloração de Gram.
Bactérias Gram-negativas:
Composta de:
- Uma ou poucas camadas de peptidoglicano;
- Espaço periplasmático: matriz concentrada parecida com gel no espaço entre a membrana citoplasmática interior e a membrana bacterial exterior; contém proteínas envolvidas no transporte, sensoriamento químico e outras funções celulares importantes;
- As porinas conferem permeabilidade através da membrana externa;
- Uma membrana externa que contém LPS (lipopolissacarídeo);
- Coram-se de rosa pela Coloração de Gram.
O citoplasma das células procarióticas mantém uma concentração elevada de solutos dissolvidos, o que cria uma significativa pressão osmótica – cerca de 2 atm (203 kPa) – em uma célula típica. Trata-se da mesma pressão em um pneu de automóvel. Para resistir a essa pressão e evitar uma explosão (lise celular), a maioria das células de bactérias e arqueias possui uma parede. Além de proteger contra a lise osmótica, as paredes celulares conferem forma e rigidez à célula.
O conhecimento da estrutura e função da parede celular é importante não apenas para o entendimento de como as células procarióticas funcionam, mas também devido ao fato de que muitos antibióticos têm como alvo a síntese da parede celular, deixando a célula suscetível à lise. Uma vez que as células humanas carecem de parede celular, esses antibióticos são obviamente vantajosos no tratamento de infecções bacterianas.
O peptideoglicano é um polissacarídeo encontrado apenas em bactérias que consiste em uma repetição alternada de N-acetilglicosamina e ácido N-acetilmurâmico, o último ligado por ligações cruzadas por meio de tetrapeptídeos em filamentos adjacentes. A enzima lisozima e o antibiótico penicilina possuem como alvo o peptideoglicano, levando à lise celular.
As paredes celulares de arqueias podem ser de diferentes tipos, incluindo pseudomureína, vários polissacarídeos e camadas S, que são compostas por proteínas ou glicoproteínas. Assim como em bactérias, as paredes de arqueias protegem a célula contra a lise osmótica.
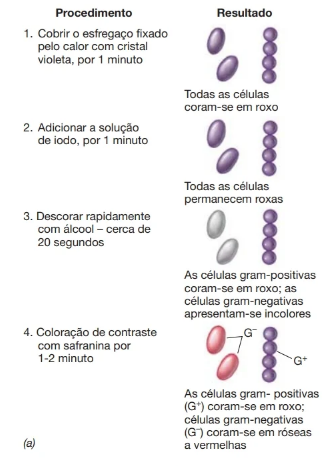
Corantes que conferem diferentes cores a diferentes tipos de células são denominados corantes diferenciais. Um importante procedimento de coloração diferencial, amplamente utilizado em microbiologia, é a coloração de Gram. De acordo com sua reação à coloração de Gram, bactérias podem ser divididas em dois grupos principais: gram-positivas e gram-negativas. Após a coloração de Gram, as bactérias gram-positivas coram-se em roxo-violeta, enquanto as bactérias gram-negativas, em cor-de-rosa. Essa diferença de reação à coloração de Gram deve-se às diferenças na estrutura da parede celular das células gram-positivas e gram-negativas. Após a coloração com um corante básico, como o cristal violeta que confere às células uma coloração roxa, o tratamento com o etanol descora as células gram-negativas, mas não as gram-positivas. Após a coloração de contraste com um corante de cor diferente, normalmente a safranina, os dois tipos de células podem ser distinguidos microscopicamente por suas cores diferenciadas.
A coloração de Gram é o procedimento de coloração mais utilizado na microbiologia, sendo frequentemente utilizado para iniciar a caracterização de uma bactéria recentemente isolada. Se houver a disponibilidade de um microscópio fluorescente, a coloração de Gram pode ser reduzida a um procedimento de uma etapa; as células gram-positivas e gram-negativas fluorescem em diferentes cores quando tratadas com um produto químico especial
Paredes celulares de bactérias. (a, b) Diagramas esquemáticos de paredes celulares gram-positivas e gram-negativas. A foto da coloração de Gram no centro mostra células de
Staphylococcus aureus (roxa, gram-positiva) e
Escherichia coli (cor-de-rosa, gram-negativa). (c, d) Micrografias eletrônicas de transmissão (MET) apresentando a parede celular de uma bactéria gram-positiva e de uma bactéria gram-negativa. (e, f) Micrografias eletrônicas de varredura de bactérias gram-positivas e gram-negativas, respectivamente. Observe as diferenças na textura da superfície das células apresentadas. Cada célula em MET tem aproximadamente 1 mm de largura.


O mecanismo da coloração de Gram se refere à composição da parede celular, sendo que as Gram-positivas possuem uma espessa camada de peptideoglicano e ácido teicóico, e as Gram-negativas, uma fina camada de peptideoglicano, sobre a qual se encontra uma camada composta por lipoproteínas, fosfolipídeos, proteínas e lipopolissacarídeos. Durante o processo de coloração, o tratamento com álcool-acetona extrai os lipídeos, daí resultando uma porosidade ou permeabilidade aumentada da parede celular das bactérias Gram-negativas.
Assim, o complexo cristal violeta-iodo (CVI) pode ser retirado e as bactérias Gram-negativas são descoradas. A parede celular das bactérias gram-positivas, em virtude de sua composição diferente, torna-se desidratada durante o tratamento com álcool-acetona, a porosidade diminui, a permeabilidade é reduzida e o complexo CVI não pode ser extraído.
Outra explicação baseia-se também em diferenças de permeabilidade entre os dois grupos de bactérias. Nas Gram-positivas, o complexo CVI é retido na parede após tratamento pelo álcool-acetona, o que causa, provavelmente, uma diminuição do diâmetro dos poros da camada de glicopeptídeo ou peptideoglicano da parede celular. A parede das bactérias Gram-negativas permanece com porosidade suficientemente grande, mesmo depois do tratamento com álcool acetona, possibilitando a extração do
complexo CV-l.


Camadas S:
As paredes celulares de arqueias podem ser de diferentes tipos, incluindo pseudomureína, vários polissacarídeos e camadas S, que são compostas por proteínas ou glicoproteínas. Assim como em bactérias, as paredes de arqueias protegem a célula contra a lise osmótica.
O tipo mais comum de parede celular em arqueias corresponde a uma camada superficial paracristalina, ou camada S, como é chamada. As camadas S consistem em moléculas entrelaçadas de proteínas ou glicoproteínas. A estrutura paracristalina das camadas S pode organizar-se em várias simetrias, como hexagonal, tetragonal ou trimérica, dependendo do número e estrutura das subunidades a partir das quais são formadas. Camadas S foram detectadas em representantes de todas as principais linhagens de arqueias, e também em diversas espécies de bactérias.
As paredes celulares de algumas arqueias, como, por exemplo, aquela do organismo metanogênico Methanocaldococcus jannaschii, consistem em somente uma camada S. Portanto, as camadas S são suficientemente resistentes para suportar pressões osmóticas sem qualquer outro tipo de componente de parede. No entanto, em muitos organismos, as camadas S encontram-se presentes em adição a outros componentes da parede celular, geralmente polissacarídeos. Por exemplo, em Bacillus brevis, uma espécie de bactéria uma camada S está presente juntamente com o peptideoglicano. Contudo, quando uma camada S está presente em conjunto com outros componentes da parede, a camada S corresponde sempre à camada mais externa da parede, aquela que está em contato direto com o ambiente.
Além de atuar na proteção contra a lise osmótica, as camadas S podem apresentar outras funções. Por exemplo, com a interface entre a célula e seu ambiente, é provável que a camada S atue como uma barreira seletiva, permitindo a passagem de solutos de baixa massa molecular e excluindo moléculas e estruturas grandes (como os vírus). A camada S pode também atuar na retenção de proteínas próximo à superfície celular, como a membrana externa o faz em bactérias gram-negativas. Portanto, podemos observar diversas estruturas de parede celular em espécies de arqueias, variando desde aquelas que se assemelham aos peptideoglicanos, até aquelas que não possuem nenhum polissacarídeo. Contudo, a não ser em raras exceções, todas as arqueias possuem algum tipo de parede celular e, como nas bactérias, a parede celular das arqueias age na prevenção da lise osmótica e no estabelecimento da forma da célula. Como as arqueias não possuem peptideoglicanos, elas são naturalmente resistentes à ação da lisozima e do antibiótico penicilina, agentes que destroem os peptideoglicanos ou que interrompem sua biossíntese
Citoplasma:
• Local onde ocorre reações químicas vitais para a célula;
• Entre elas a fabricação de moléculas que irão constituir as estruturas celulares. Também é responsável pelo armazenamento de substâncias de reserva.
Ribossomos:
• Local de síntese de proteínas e enzimas. Alvo importante para drogas antibacterianas.

Os ribossomos correspondem aos sítios da síntese proteica. Uma célula pode conter vários milhares de ribossomos, sendo esse número relacionado diretamente à taxa de crescimento. Cada ribossomo é composto por duas subunidades. Os procariotos apresentam as subunidades ribossomais 30S e 50S, originando ribossomos intactos 70S. Os eucariotos apresentam as subunidades ribossomais 40S e 60S, originando ribossomos intactos 80S. Os valores S correspondem a unidades Svedberg, que se referem ao coeficiente de sedimentação das subunidades ribossomais (30S e 50S nos procariotos, 40S e 60S nos eucariotos) ou de ribossomos intactos (70S nos procariotos e 80S nos eucariotos), quando submetidos à força centrífuga em uma ultracentrífuga. (Embora partículas maiores exibam valores S maiores, a relação não é linear e, dessa forma, os valores S não podem ser simplesmente somados.)
Cada subunidade ribossomal contém RNA ribossomais e proteínas ribossomais específicas. A subunidade 30S contém o RNAr 16S e 21 proteínas, enquanto a subunidade 50S contém os RNArs 5S e 23S, além de 31 proteínas. Assim, em Escherichia coli, são encontradas 52 proteínas ribossomais diferentes, a maioria presente como cópia única por ribossomo. O ribossomo é uma estrutura dinâmica, cujas subunidades associam-se e dissociam-se alternadamente e também interagem com várias outras proteínas. Diversas proteínas essenciais à atividade ribossomal interagem com o ribossomo em vários estágios da tradução. Essas são consideradas “fatores de tradução”, em vez de “proteínas ribossomais” por si só.
Fissão binária:
Em microbiologia, o crescimento é definido como um aumento no número de células. As células microbianas possuem tempo de vida limitado, e uma espécie é mantida apenas como resultado do crescimento contínuo de sua população. À medida que as macromoléculas acumulam-se no citoplasma de uma célula, elas são agrupadas em importantes estruturas, como parede celular, membrana citoplasmática, flagelos, ribossomos, complexos enzimáticos e assim por diante, eventualmente levando à divisão celular.
Em uma cultura em crescimento de uma bactéria em forma de bastonete, como Escherichia coli, as células alongam-se até aproximadamente duas vezes o seu tamanho original e, então, formam uma partição que divide a célula em duas células-filhas. Esse processo é chamado de fissão binária (“binário” expressa o fato de duas células originarem-se a partir de uma). Essa partição é chamada de septo e resulta de uma invaginação da membrana citoplasmática e da parede celular de direções opostas; a formação do septo continua até a individualização das duas células-filhas. Há algumas variações no padrão geral da fissão binária. Em algumas bactérias, como Bacillus subtilis, o septo forma-se sem a constrição da parede celular, ao passo que no brotamento de Caulobacter a constrição ocorre, mas nenhum septo é formado. Entretanto, em todos os casos, quando uma célula eventualmente separa-se para formar duas células, dizemos que ocorreu uma geração, e o tempo requerido para esse processo é chamado de tempo de geração.
Durante uma geração, todos os constituintes celulares aumentam proporcionalmente e as células estão em um crescimento balanceado. Cada célula-filha recebe um cromossomo e cópias suficientes de ribossomos e todos os outros complexos macromoleculares, monômeros e íons inorgânicos para existir como uma célula independente. A partição da molécula de DNA replicada entre as duas células-filhas depende de o ancoramento do DNA à membrana citoplasmática ser mantido durante a divisão, com a constrição levando à separação dos cromossomos, um para cada célula-filha
Flagelo:
• Função: confere movimento à célula (taxia): velocidade 200 – 500 μm/segundo;
• Composto por: uma estrutura basal, um gancho e um longo filamento de FLAGELINA.
A motilidade de natação acontece devido à presença dos flagelos. O flagelo é composto por diversas proteínas e é ancorado na parede celular e na membrana citoplasmática. Em bactérias, o filamento flagelar é constituído da proteína flagelina e rotaciona com a energia fornecida pela força próton-motiva. O flagelo de arqueias e bactérias difere na estrutura e na forma como a energia é acoplada à rotação.
Os flagelos de arqueias são significativamente mais delgados que os flagelos bacterianos, exibindo somente 10 a 13 nm de espessura, porém conferem movimento de rotação à célula como observado em bactérias. De modo diverso ao observado em bactérias, onde há uma única proteína no filamento flagelar, várias flagelinas diferentes são encontradas em arqueias sendo que as sequências de aminoácidos e os genes que codificam as flagelinas de arqueias não exibem qualquer relação com aquelas da flagelina bacteriana. O diâmetro significativamente menor do flagelo de arqueias, comparado ao flagelo bacteriano, possivelmente reduz o torque e, consequentemente, a potência do motor flagelar, de modo que velocidades natatórias menores não seriam surpreendentes. Além disso, a partir de experimentos bioquímicos com Halobacterium, é possível inferir que os flagelos de arqueias são alimentados diretamente por ATP e não pela força próton-motiva, a fonte de energia dos flagelos de bactérias .
Um flagelo consiste em vários componentes e movimenta-se por rotação, muito similar à hélice de um barco a motor. A base do flagelo é estruturalmente distinta do filamento. A base do flagelo apresenta uma região mais larga, denominada gancho. O gancho é composto por um único tipo de proteína e conecta o filamento à porção motora do flagelo na base. A porção motora do flagelo é ancorada na membrana citoplasmática e na parede celular. O motor é formado por um bastão central que passa por meio de uma série de anéis. Em bactérias gram-negativas, um anel externo, chamado de anel L, é ancorado na camada lipopolissacarídica. Um segundo anel, chamado de anel P, é ancorado na camada de peptideoglicano da parede celular. Um terceiro conjunto de anéis, chamados de MS e anéis C, está localizado dentro da membrana citoplasmática e do citoplasma, respectivamente (Figura 2.51a). Em bactérias gram-positivas, que não possuem membrana externa, apenas o par interior de anéis se mostra presente. Ao redor do anel interior e ancorado na membrana citoplasmática, está uma série de proteínas denominadas proteínas Mot. Um conjunto final de proteínas, denominadas proteínas Fli (Figura 2.51a), funciona como o interruptor do motor, invertendo a direção da rotação dos flagelos em resposta a sinais intracelulares.

As bactérias que se movem por deslizamento não utilizam a rotação dos flagelos, mas, em vez disso, se arrastam ao longo de uma superfície sólida empregando um dos diferentes mecanismos, incluindo excreção polissacarídica, pulsação ou proteínas de deslizamento rotativo.
Bactérias móveis respondem a gradientes químicos e físicos em seus ambientes, controlando o comprimento das corridas e a frequência das oscilações. As oscilações são controladas pela direção de rotação do flagelo, que, por sua vez, é controlada por uma rede de proteínas sensoriais e de resposta.
Pili e fímbrias:
Função:
- Pili sexual;
- Sítios aceptores de vírus;
- Adesão à células receptoras de mamíferos.
As fímbrias e os pili são proteínas filamentosas que se projetam a partir da superfície de uma célula, podendo apresentar muitas funções, incluindo ligação, intercâmbio genético e motilidade pulsante.
As fímbrias conferem às células a capacidade de adesão a superfícies, incluindo tecidos animais no caso de algumas bactérias patogênicas, ou de formação de películas (camadas delgadas de células sobre uma superfície líquida) ou biofilmes em superfícies sólidas. Patógenos humanos notórios nos quais essas estruturas auxiliam no processo da doença incluem espécies de Salmonella (salmonelose), Neisseria gonorrhoeae (gonorreia) e Bordetella pertussis (coqueluche).
Os pili assemelham-se às fímbrias, porém são estruturas normalmente mais longas, estando presentes na superfície celular em uma ou poucas cópias. Como os pili podem ser receptores de determinados tipos de vírus, tais estruturas podem ser mais bem visualizadas ao microscópio eletrônico quando se encontram revestidas por partículas virais. São conhecidas várias classes de pili, diferenciadas por sua estrutura e função. Duas funções bastante importantes dos pili incluem facilitar a troca genética entre as células em um processo chamado de conjugação, e impedir a adesão de patógenos a tecidos específicos do hospedeiro que eles venham a invadir. A última função tem sido bem estudada em patógenos gram-negativos como as Neisseria, espécies que causam a gonorreia e a meningite, porém os pili também estão presentes em determinados patógenos gram-positivos como Streptococcus pyogenes, a bactéria que causa a faringite estreptocócica e a escarlatina.
Uma classe importante de pili, denominada pili tipo IV, auxilia a adesão celular, mas também permite uma forma incomum de motilidade celular, denominada motilidade pulsante. Os pili tipo IV estão presentes apenas nos polos dessas células em forma de bastonete que os contêm. A motilidade pulsante é um tipo de motilidade por deslizamento, um movimento ao longo de uma superfície sólida (Seção 2.18). Na motilidade pulsante, a extensão dos pili, seguida de sua retração, arrasta a célula ao longo de uma superfície sólida, com energia proveniente do ATP. Determinadas espécies de Pseudomonas e Moraxella são bem conhecidas por sua motilidade pulsante.
Os pili tipo IV também foram implicados como fatores essenciais de colonização para determinados patógenos de seres humanos, incluindo Vibrio cholerae (cólera) e Neisseria gonorrhoeae (gonorreia). Possivelmente, a motilidade pulsante desses patógenos auxilia os organismos a localizarem sítios específicos para a adesão, para então dar início ao processo de doença. Acredita-se que os pili tipo IV também atuem como mediadores na transferência genética, por meio do processo de transformação em algumas bactérias, o qual, juntamente com a conjugação e a transdução, são os três meios mais conhecidos de transferência gênica horizontal em procariotos.


Inclusões celulares:
Inclusões estão frequentemente presentes em células procarióticas. As inclusões funcionam como reservas de energia e/ou como reservatórios de carbono, ou desempenham funções especiais. As inclusões podem ser visualizadas diretamente ao microscópio óptico, sendo normalmente envoltas por membranas de camada única (sem unidade) que as isolam no interior da célula. O armazenamento de carbono ou de outras substâncias em uma forma insolúvel é vantajoso para a célula, pois reduz o estresse osmótico que existiria caso a mesma quantidade da substância fosse dissolvida no citoplasma.
Células procarióticas contêm inclusões de enxofre, polifosfato, polímeros de carbono ou minerais que formam partículas magnéticas (magnetossomos). Estas substâncias funcionam como materiais de armazenamento ou na magnetotaxia.
Polímeros de armazenamento de carbono:
Um dos corpos de inclusão mais comuns em organismos procarióticos corresponde ao ácido poli-β-hidroxibutírico (PHB), um lipídeo formado por unidades de ácido β-hidroxibutírico. Os monômeros de PHB polimerizam-se por ligações do tipo éster, e em seguida o polímero se agrega em grânulos; estes últimos podem ser vistos por microscopia óptica ou eletrônica.
O monômero no polímero normalmente é o hidroxibutirato (C4), contudo o comprimento desse monômero pode variar consideravelmente, desde C3 até C18. Dessa forma, o termo mais genérico, poli-β-hidroxialcanoato (PHA), é geralmente empregado para descrever essa classe de polímeros de armazenamento de carbono e energia. PHAs são sintetizados pelas células quando há um excesso de carbono, e são clivados para uso como fontes de carbono ou energia quando as condições o permitem. Muitas bactérias e arqueias produzem PHAs.
Outro produto de armazenamento é o glicogênio, que corresponde a um polímero de glicose e, assim como os PHAs, é um reservatório de carbono e energia, produzido quando há excesso de carbono no ambiente.
Polifosfato:
Muitos microrganismos acumulam fosfato inorgânico (PO43-) na forma de grânulos de polifosfato. Esses grânulos podem ser degradados e utilizados como fontes de fosfato na biossíntese de ácidos nucleicos e fosfolipídeos e, em alguns organismos, podem ser utilizados diretamente na produção do composto rico em energia, ATP. Frequentemente, o fosfato é um nutriente limitante em ambientes naturais. Portanto, caso uma célula encontre-se e em uma situação em que há excesso de fosfato, é vantajoso o armazenamento deste na forma de polifosfato para uso futuro.
Enxofre:
Muitos procariotos gram-negativos são capazes de oxidar compostos sulfurados reduzidos, como o sulfeto de hidrogênio (H2S); esses organismos correspondem às “bactérias sulfurosas”, descobertas pelo grande microbiologista Sergei Winogradsky. A oxidação de sulfeto está associada à necessidade de elétrons para conduzir as reações do metabolismo energético (quimiolitotrofia) ou de fixação de CO2
(autotrofia).
Em ambos os casos, pode haver o acúmulo de enxofre elementar (S⁰) nas células, proveniente da oxidação do sulfeto, em grânulos visíveis microscopicamente. Esses glóbulos de enxofre elementar permanecem enquanto a fonte de enxofre reduzido de onde eles são derivados estiver presente. Entretanto, quando a fonte de enxofre reduzido torna-se limitante, o enxofre presente nos grânulos é oxidado a sulfato (SO42-,), com o desaparecimento gradual dos grânulos à medida que essa reação ocorre. Interessantemente, embora os glóbulos de enxofre aparentemente residam no citoplasma, eles estão presentes, na realidade, no periplasma. Nessas células, o periplasma expande-se a fim de acomodar os glóbulos, à medida que H2S é oxidado a S⁰, e então contrai-se à medida que S⁰ é oxidado a SO42-.

Minerais carbonatos:
Cianobactérias filamentosas são conhecidas por formar minerais carbonatos na superfície externa de suas células. No entanto, algumas cianobactérias também formam minerais carbonatos dentro das células, como inclusões celulares. Por exemplo, a cianobactéria unicelular Gleomargarita forma grânulos intracelulares de bentonita, um mineral carbonato que contém bário, estrôncio e magnésio.
O processo microbiológico de formação de minerais é chamado de biomineralização. Ainda não está claro exatamente o porquê desse mineral em particular ser formado por essa cianobactéria, embora possa servir como um fator de resistência para a manutenção das células em seu hábitat, no fundo de um lago alcalino no México. A biomineralização de diversos minerais diferentes é catalisada por vários procariotos, contudo apenas no caso da Gleomargarita e dos magnetossomos (que serão discutidos a seguir) é que há o desenvolvimento de inclusões intracelulares.
Inclusões magnéticas de armazenamento: magnetossomos:
Algumas bactérias são capazes de orientar-se especificamente em um campo magnético pelo fato de conterem magnetossomos. Essas estruturas são partículas intracelulares compostas pelo mineral óxido de ferro magnetita – Fe3O4. Em algumas bactérias magnetotáticas é formado o mineral contendo enxofre greigite (Fe3S4). Tanto a magnetita quanto o greigite são minerais magnetotáticos.
Os magnetossomos geram um dipolo magnético na célula, permitindo-a orientar-se em um campo magnético. Bactérias que produzem magnetossomos exibem magnetotaxia, um processo de orientação ao longo das linhas do campo magnético da Terra.
Os magnetossomos foram encontrados em vários organismos aquáticos, os quais, em culturas laboratoriais, apresentam melhor crescimento em baixas concentrações de O2. Assim, surgiu a hipótese de que uma das funções dos magnetossomos seja guiar essas células aquáticas para baixo (a direção do campo magnético da Terra) em direção aos sedimentos onde as concentrações de O2 são menores. Um produtor de greigite é uma bactéria redutora de sulfato, e esses organismos são anaeróbios obrigatórios. Permanecer em zonas anóxicas seria especialmente importante para essas espécies magnetotáticas
Um magnetossomo individual é envolto por uma fina membrana composta por fosfolipídeos, proteínas e glicoproteínas. Embora essa membrana não corresponda a uma unidade de membrana real (bicamada), como a membrana citoplasmática, as proteínas da membrana do magnetossomo são funcionais, uma vez que catalisam a precipitação de Fe3+ no magnetossomo em desenvolvimento. Uma
membrana similar também circunda os grânulos de PHA e os glóbulos de enxofre. A morfologia dos magnetossomos parece ser espécie-específica, variando em forma desde quadrada a retangular até afilada. Nenhuma arqueia contendo magnetossomos foi descoberta até o momento.
Referências
MADIGAN, Michael T.; MARTINKO, John M.; BENDER, Kelly S.; et al. Microbiologia de Brock. Grupo A, 2016. E-book. ISBN 9788582712986. Disponível em: https://app.minhabiblioteca.com.br/#/books/9788582712986/. Acesso em: 07 abr. 2024.














































{kind=link}





Nenhum comentário:
Postar um comentário