Olá pessoal, hoje eu irei trazer mais um tema em microbiologia: apontamentos sobre nutrição, crescimento e cultivo de microrganismos - sobretudo bactérias.
Nutrição microbiana:
• Metabolismo (conjunto de todas as transformações químicas de uma célula ou de um organismo) = catabolismo (fase de degradação do metabolismo) + anabolismo (biossíntese);
• Macronutrientes (necessários em grandes quantidades) e micronutrientes (necessários em quantidades traço).
Macronutrientes:
• Carbono (C): 50% do peso seco da célula; aminoácidos, ácidos graxos, açúcares, bases nitrogenadas etc.;
• Nitrogênio (N): 12% do peso seco da célula; proteínas, ácidos nucleicos etc., maior parte na forma inorgânica amônia (NH3), nitrato (NO3-), gás nitrogênio (N2);
• Fósforo (P): encontrado na natureza como fosfatos orgânicos e inorgânicos; compõe ácidos nucleicos, fosfolipídios etc.;
• Enxofre (S): mais encontrado em fontes inorgânicas, sulfato (SO₄²-) e sulfeto ([HS]−) - compõe aminoácidos cisteína e metionina, vitaminas, coenzima A etc.;
• Potássio (K): compõe enzimas envolvidas na síntese proteica;
• Magnésio (Mg): estabiliza ribossomos, membranas, ácidos nucleicos; cofator de enzimas;
• Cálcio (Ca): estabiliza parede celular e membrana citoplasmática; termoestabilidade de endósporos;
• Ferro (Fe): importante na respiração celular, componente de citocromos e outras proteínas envolvidas nas reações de transporte de elétrons; condições anóxicas -> forma solúvel, ferro ferroso (Fe2+); condições óxicas -> forma insolúvel, ferro férrico (Fe³⁺); sideróforos -> agentes ligantes de ferro -> ácido hidroxâmico.
Micronutrientes:
• Elementos traço: compõe enzimas;
• Fatores de crescimento: compostos orgânicos necessários em pequenas quantidades - vitaminas, aminoácidos, purinas e pirimidinas.
Cultivo microbiano:
• Meios definidos: são preparados pela adição de quantidades precisas de compostos químicos inorgânicos ou orgânicos à água destilada. Portanto, a composição química exata de um meio definido (de forma qualitativa e quantitativa) é conhecida. A fonte de carbono é de importância fundamental em qualquer meio de cultura, uma vez que todas as células necessitam de grandes quantidades de carbono a fim de sintetizarem novo material celular. A natureza da fonte de carbono e sua concentração dependem do organismo a ser cultivado.
• Meios complexos: para o cultivo de muitos microrganismos, o conhecimento da composição exata de um meio não é essencial. Nessas situações, meios complexos podem ser suficientes ou até mesmo vantajosos. Os meios complexos empregam componentes de produtos microbianos, animais ou vegetais, como a caseína (proteína do leite), carne (extrato de carne), soja (caldo tríptico de soja), células de leveduras (extrato de levedura), ou várias outras substâncias altamente nutritivas. Esses produtos de digestão encontram-se disponíveis comercialmente na forma desidratada, e necessitam apenas de hidratação para originar um meio de cultura. Entretanto, a desvantagem no uso de um meio complexo consiste no fato de que a
sua composição nutricional não é precisamente conhecida.
• Meios enriquecidos: um meio enriquecido, utilizado na cultura de microrganismos nutricionalmente exigentes (fastidiosos), muitos dos quais são patógenos (nutrientes ''mimetizam'' condições do hospedeiro), começa como um meio complexo e, em seguida, é suplementado com substâncias adicionais altamente nutritivas como o soro ou sangue.
Em situações particulares, especialmente na microbiologia diagnóstica, frequentemente os meios de cultura são produzidos de modo a tornarem-se seletivos ou diferenciais (ou ambos).
• Meios seletivos: contém compostos que inibem seletivamente o crescimento de alguns microrganismos, mas não o de outros. Por exemplo, meios seletivos estão disponíveis para o isolamento de determinados patógenos, como linhagens de Salmonella ou Escherichia coli que causam intoxicação alimentar.
• Meios diferenciais: corresponde àquele ao qual um indicador, normalmente um corante, é adicionado, revelando por meio de uma mudança de coloração se uma reação metabólica em particular ocorreu durante o crescimento. Os meios diferenciais são bastante úteis na distinção de espécies bacterianas, e são amplamente utilizados no diagnóstico clínico e na microbiologia sistemática.
Cultivo em laboratório:
• Meio estéril -> cultura pura;
• Meio líquido X meio sólido (colônias);
• Uso de técnicas assépticas.
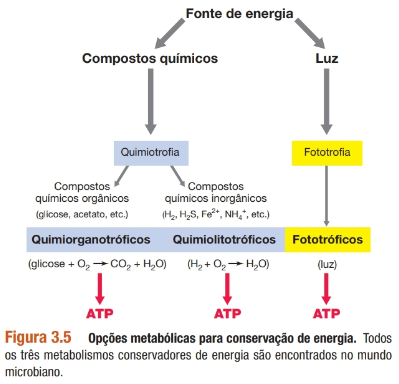
Classes energéticas dos microrganismos:
A energia conservada é retida na célula na forma de ligações ricas em energia do composto trifosfato de adenosina (ATP).
• Quimiotróficos: organismos que conservam energia a partir de compostos químicos;
• Quimiorganotróficos: organismos que conservam energia a partir de compostos químicos orgânicos;
- Aeróbios: alguns microrganismos podem obter energia a partir de compostos orgânicos somente na presença de oxigênio;
- Anaeróbios: outros extraem a energia somente na ausência de oxigênio;
- Anaeróbios facultativos: há ainda outros que podem metabolizar compostos orgânicos tanto na presença quanto na ausência de oxigênio.
• Quimiolitotróficos: organismos que conservam energia a partir de compostos químicos inorgânicos;
Diversos compostos inorgânicos podem ser oxidados, como, por exemplo, H₂, H₂S (sulfeto de hidrogênio), NH3 (amônia) e Fe2+ (ferro ferroso). Grupos próximos de quimiolitotróficos normalmente se especializam na oxidação de grupos relacionados de compostos inorgânicos, e, dessa forma, há a bactéria “sulfurosa”, a bactéria do “ferro”, a bactéria “nitrificante”, e assim por diante.
• Fototróficos: microrganismos fototróficos contêm pigmentos que os permitem converter a energia luminosa em energia química e, portanto, diferentemente dos quimiotróficos, não necessitam de compostos químicos como uma fonte de energia.
- Fotossíntese oxigênica: há produção de oxigênio (O₂). Entre os microrganismos, a fotossíntese oxigênica é característica de cianobactérias, que são procariotos, e algas, que são eucariotos;
- Fotossíntese anoxigênica: realizada por bactérias púrpuras e verdes e pelas heliobactérias (todas as bactérias), e não resulta na produção de O₂.
Ambos os grupos de fototróficos apresentam grandes semelhanças em seus mecanismos de síntese de ATP, resultado do fato de que a fotossíntese oxigênica evoluiu da forma anoxigênica mais simples há cerca de 3 bilhões de anos.
Independente de como um microrganismo conserva a energia, temos visto que todas as células necessitam de grandes quantidades de carbono para a produção de novos materiais celulares.
• Heterotróficos: se um organismo é heterotrófico, o carbono celular é obtido a partir de algum composto químico orgânico; por definição, os organismos quimiorganotróficos são também heterotróficos;
• Autotróficos: utiliza dióxido de carbono (CO₂) como sua fonte de carbono. Denominados produtores primários, uma vez que sintetizam nova matéria orgânica a partir de CO₂. Praticamente toda a matéria orgânica presente na Terra foi sintetizada por produtores primários, particularmente, por fototróficos.
Crescimento microbiano:
• Crescimento = aumento na população (número de células);
• Fissão binária em E. coli: elongação até o dobro do tamanho
Síntese do peptidoglicano/mureína:
• Autolisinas: hidrolisam ligações β1->4 de NAG-NAM -> novo material é adicionado nas ''aberturas'' -> amilase salivar;
• Bactoprenol: transporta precursores do peptidoglicano através da membrana citoplasmática -> interage com glicolases, que inserem os precursores no local de crescimento -> bacitracina;
• Transpeptidação: ocorre exatamente à membrana plasmática, onde não há ATP disponível -> penicilina e vancomicina.
A síntese do novo peptideoglicano durante o crescimento envolve a clivagem controlada do peptideoglicano preexistente, com a inserção simultânea dos precursores de peptideoglicano.
O bactoprenol transporta os precursores do peptideoglicano através da membrana citoplasmática, tornando-os suficientemente hidrofóbicos para atravessarem o interior da membrana. Uma vez no periplasma, o bactoprenol interage com enzimas denominadas transglicolases, que inserem os precursores no ponto de crescimento da parede celular e catalisam a formação da ligação glicosídica.
Previamente, as pequenas aberturas existentes no peptideoglicano são feitas por enzimas denominadas autolisinas, as quais têm a função de hidrolisar as ligações que conectam a N-acetilglicosamina e o ácido N-acetilmurâmico no esqueleto do peptideoglicano. O novo material da parede celular é, então, adicionado por meio dessas aberturas.
Novas unidades do tetrapeptídeo devem ser alocadas no peptideoglicano existente imediatamente após a atividade da autolisina, a fim de evitar uma ruptura na integridade do peptideoglicano no ponto de junção; uma quebra poderia causar a lise celular espontânea, chamada de autólise.
A etapa final na síntese da parede celular consiste na transpeptidação. A transpeptidação forma as ligações peptídicas cruzadas entre os resíduos de ácido murâmico em cadeias adjacentes de glicano.
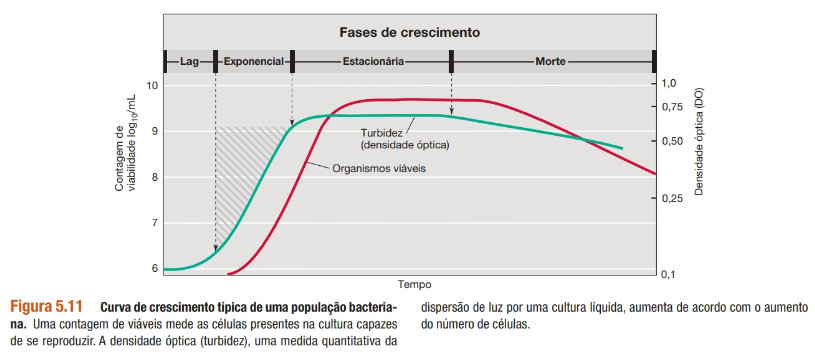
Crescimento microbiano: crescimento exponencial.
As células microbianas sofrem crescimento exponencial, e um gráfico semilogarítmico do número de células pelo tempo pode revelar o tempo de duplicação da população. Matemática simples pode ser usada para calcular várias expressões de crescimento a partir de dados de números de células. Expressões-chave são n, o número de gerações; t, tempo; e g, tempo de geração. O tempo de geração é expresso como g = t/n.
Este padrão de aumento populacional, em que o número de células é duplicado a um intervalo de tempo constante, é denominado crescimento exponencial.
Em um experimento desse tipo, quando o número de células é plotado em um gráfico de coordenadas aritméticas (lineares) em função do tempo, obtém-se uma curva de inclinação constantemente crescente.
Em contrapartida, quando o número de células é plotado em uma escala logarítmica em função do tempo (um gráfico semilogarítmico), obtém-se uma linha reta. Essa função linear reflete o fato de que as células estão crescendo exponencialmente e a população está se duplicando em um intervalo de tempo constante. Gráficos semilogarítmicos também são convenientes para estimar os tempos de geração de uma cultura a partir de um conjunto de dados de crescimento.

Há uma relação fixa entre o número de células inicialmente presentes em uma cultura e o número presente após um período de crescimento exponencial; essa relação pode ser expressa como
N = N0 * 2^n
em que:
N: número final de células
N0: número inicial de células
n: número de gerações formadas durante o período de crescimento exponencial.
O tempo de geração (g) da população em crescimento exponencial corresponde a
g = t/n
em que t refere-se à duração do crescimento exponencial, sendo expresso em dias, horas ou minutos. A partir do conhecimento dos números inicial e final de células em uma população em crescimento exponencial, é possível calcular n e, a partir de n e do conhecimento de t, o tempo de geração, g.
Principais fases do crescimento microbiano:
• Fase lag ou de adaptação: – praticamente não há aumento no número de células. É uma fase de intenso metabolismo onde as células individuais aumentam de tamanho. Células fisiologicamente ativas e sintetizando novas enzimas para se adaptarem ao novo meio. O período de tempo em que permanecem nesta fase é dependente da cepa;
• Fase log ou exponencial: – o crescimento é máximo em taxas exponenciais. Alta atividade metabólica com células aproximadamente uniformes em termos de composição química e atividades metabólicas e fisiológicas. Pico da atividade e eficiência metabólica. Fase de interesse industrial, porém, os microrganismos podem estar mais suscetíveis a antimicrobianos;
• Fase estacionária: – o número de indivíduos novos é equivalente aos mortos, não é mais observado crescimento. Há acúmulo de produtos metabólicos tóxicos e/ou exaustão de nutrientes;
• Fase de declínio ou morte: – acúmulo adicional de produtos metabólicos inibitórios e depleção dos nutrientes essenciais. A taxa de morte é acelerada e o número de células viáveis diminui de forma exponencial. Dependendo da espécie, pode haver destruição da cultura ou restarem algumas células vivas que resistirão até o final da curva. Tipicamente, todas as células normalmente morrem em dias a meses.
Contagem de células viáveis:
Uma célula viável é definida como aquela capaz de dividir-se, originando células-filhas, e, na maioria das situações de contagens de células, são as células que causam maior interesse. Com esse propósito, podemos utilizar um método de contagem de células viáveis, também denominado contagem em placa, uma vez que placas de ágar são requeridas. O procedimento de contagem de células viáveis pressupõe que cada célula viável é capaz de crescer e dividir-se, originando uma colônia. Assim, o número de colônias e o número de células são proporcionais.
No método de semeadura por espalhamento, um volume (geralmente 0,1 mL ou menos) da cultura diluída apropriadamente é espalhado sobre a superfície de um meio sólido com auxílio de uma alça de vidro estéril.
No método de semeadura em profundidade, um volume conhecido (geralmente de 0,1–1,0 mL) da cultura é pipetado em uma placa de Petri estéril. O meio de cultura fundido é, então, adicionado e bem misturado, movimentando-se delicadamente a placa em círculos sobre a bancada.
Tanto no método de semeadura por espalhamento quanto na semeadura em profundidade, é importante que o número de colônias desenvolvidas na placa não seja muito elevado, nem muito baixo. Em placas muito populosas, algumas células podem não formar colônias e algumas colônias podem se fundir, levando a erros de contagem. Se o número de colônias for muito pequeno, a significância estatística da contagem será baixa. A prática geral, de maior valia estatística, consiste na contagem somente de placas que possuam entre 30 e 300 colônias.
Para que se obtenha o número apropriado de colônias, a amostra a ser contada deve quase sempre ser diluída. Tendo em vista que raramente podemos prever o número aproximado de células viáveis, em geral mais de uma diluição é necessária. Frequentemente, são empregadas várias diluições decimais da amostra. No caso de culturas densas, essas diluições seriadas são necessárias para obter-se a diluição adequada para plaquear um número contável de colônias.
Efeitos ambientais no crescimento microbiano:
• Temperatura;
• pH;
• Água livre e osmolaridade;
• Oxigenação.
Temperatura:
Para cada microrganismo há uma temperatura mínima, abaixo da qual o crescimento não é possível, uma temperatura ótima, na qual o crescimento ocorre rapidamente, e uma temperatura máxima, acima da qual o crescimento torna-se impossível. Essas três temperaturas, denominadas temperaturas cardeais.
Temperaturas frias:
• Psicrófilo → T ótima ≤ 15 °C;
• Psicrotolerante → 20 °C ≤ T ótima ≤ 40 °C, porém crescem a 0 °C;
• Adaptações à psicrofilia:
- Proteínas -> + α-hélice e - folhas β-pregueadas; + aminoácidos polares e - aminoácidos hidrofóbicos -> conferem + flexibilidade;
- Membrana citoplasmática com maior proporção de ácidos graxos
insaturados, mantendo o estado “semifluido”.
Temperaturas quentes:
• Termófilo → T ótima ≥ 45 °C;
• Hipertermófilo → T ótima ≥ 80 °C.
• Adaptações à (hiper)termofilia:
- Proteínas -> aminoácidos específicos que permitem denso ''empacotamento'' da estrutura -> termoestabilidade; + ligações iônicas entre os aminoácidos;
- Membrana citoplasmática com maior proporção de ácidos graxos
saturados (ambiente “hidrofóbico”); ou hidrocarbonetos C40 em
Archaea (monocamada lipídica).
- DNA -> presença de sais no citoplasma (evitam desnaturação); “DNA girase reversa” → superenovelamento positivo
proteína Sac7d aumenta 40 °C na T desnaturação
pH:
• pHótimo varia 2-3 unidades;
• pH mais comuns = 5 a 9;
• Poucas spp. sobrevivem em
pH < 2 ou > 10;
• “pH ótimo” refere-se ao pH do
meio externo → pH
intracelular deve ser próximo
de 7, se não, destruição de
macromolélulas.
Acidófilos:
• pH ótimo ≤ 6;
• Fungi são mais tolerantes que Bacteria → pHótimo ≤ 5;
• Acidófilos obrigatórios: Acidithiobacillus (Bacteria); Sulfolobus (Arquea) e
Thermoplasma (Arquea); Picrophilus oshimae (Arquea) – pH ótimo 0,7;
• Membrana citoplasmática estável devido ↑[H+] → em pH 7,
sofrem lise;
• Locais: solos quentes e ácidos, associados à atividade
vulcânica.
Alcalifílicos:
• pHótimo ≥ 9;
• Maioria do Domínio Archaea;
• Locais: lagos e solos ricos em carbonatos;
• Exemplo: Bacillus firmus (Bacteria);
• Como manter Força Próton Motiva se fora tem ↓[H+]? FNaM -> uma estratégia para contornar este problema em B. firmus é o uso de sódio (Na+), em vez de H+ , para conduzir as reações de transporte e a motilidade; ou seja, uma força sódio-motiva em vez de uma força próton-motiva. Curiosamente, no entanto, uma força próton motiva também é estabelecida em células de B. firmus para a síntese de ATP, embora a superfície da membrana externa seja altamente alcalina.
Osmolaridade:
• Atividade de água (aW) → ÁGUA LIVRE para utilizar em reações
bioquímicas;
• Osmose → H₂O difunde-se de regiões de alta para baixa concentração de solutos → citoplasma em “Equilíbrio aquoso positivo”;
• Água mar → 3% NaCl e + outros minerais;
• Halófilos ≠ Osmófilos ≠ Xerófilos:
- Halófilo discreto: requerem baixas concentrações de sal, entre 1% a 6%, para desenvolver;
- Halófilo moderado: requerem entre 7,5% a 15% para desenvolver;
- Halófilo extremo: dependentes de elevadas concentrações de sal para desenvolver. Em geral, > 10% de sal (15%-30%);↑ da concentração de solutos internos por bombeamento de íons inorgânicos, ou produção de solutos orgânicos;
- Osmófilo: capazes de desenvolverem em ambientes com elevada concentração de açúcares;
- Xerófilo: capazes de desenvolverem em ambientes secos diante da escassez de água.
• Em meio com solutos equivalentes a 3% NaCl → citoplasma
desidrata;
Oxigenação:
• Habitats anóxicos (livre de O
₂) → lodos, pântanos, esgotos,
regiões subterrâneas, trato gastrointestinal de animais etc.;
• Ar atmosférico → 21% de O₂;
• Gás oxigênio (O₂): diante da letalidade ou não desse gás, as bactérias podem ser classificada em:
- Aeróbios: necessitam de oxigênio para o crescimento e podem crescer em uma atmosfera padrão de 21% de oxigênio (exemplos: Mycobacterium tuberculosis, Pseudomonas aeruginosa);
- Anaeróbios facultativos: esses são aeróbios, mas na ausência de oxigênio podem realizar fermentação (exemplos: Escherichia coli e Staphylococcus aureus);
- Anaeróbios obrigatórios: Esse grupo não pode produzir energia na presença de oxigênio e são mortos pelo oxigênio (Exemplo: Clostridium botulinum);
- Microaerófilos: esse grupo é formado por aeróbios que precisam do oxigênio para produção de energia, porém não suportam os níveis de oxigênio (21%). (Exemplo: Campylobacter jejuni).
Por que os microrganismos anaeróbios são inibidos em seu crescimento ou mesmo mortos pelo oxigênio?
• Oxigênio reativo (¹O₂ , O₂ -
, H₂O₂ , OH•): promovem
oxidação espontânea de compostos celulares;
• Oxigênio singleto (¹O₂): produzido pelo
próprio organismo como
produto de reações foto
e bioquímicas.
Oxigênio molecular (O₂), não é tóxico, mas O₂ pode ser convertido em subprodutos tóxicos de oxigênio, e são eles que podem danificar ou matar as células que não são capazes de lidar com eles. Estes incluem o ânion superóxido (O₂ -), o peróxido de hidrogênio (H₂O₂) e o radical hidroxila (OH•). Todos são subprodutos da redução de O₂ a H₂O na respiração. Flavoproteínas, quinonas e proteínas de ferro-enxofre, transportadores de elétrons encontrados em praticamente todas as células, também catalisam algumas dessas reduções. Assim, independentemente de se ele pode respirar O₂, um organismo exposto ao O₂ vai experimentar formas tóxicas de oxigênio, e se não forem destruídas, essas moléculas podem causar danos nas células. Por exemplo, o ânion superóxido e OH• são fortes agentes oxidantes que podem oxidar macromoléculas e quaisquer outros compostos orgânicos na célula. Peróxidos, como H₂O₂, também podem danificar os componentes celulares, mas não são tão tóxicos quanto O₂ - ou OH•. Portanto, deve estar claro que um dos principais requisitos para habitar um mundo rico em O₂ é manter as moléculas de oxigênio tóxico sob controle.

Superóxido dismutase e outras enzimas que destroem oxigênio tóxico: adaptações para reduzir o efeito das moléculas de oxigênio tóxico.
Referências
MADIGAN, Michael T.; MARTINKO, John M.; BENDER, Kelly S.; et al. Microbiologia de Brock. Grupo A, 2016. E-book. ISBN 9788582712986. Disponível em: https://app.minhabiblioteca.com.br/#/books/9788582712986/. Acesso em: 11 abr. 2024.






































Nenhum comentário:
Postar um comentário