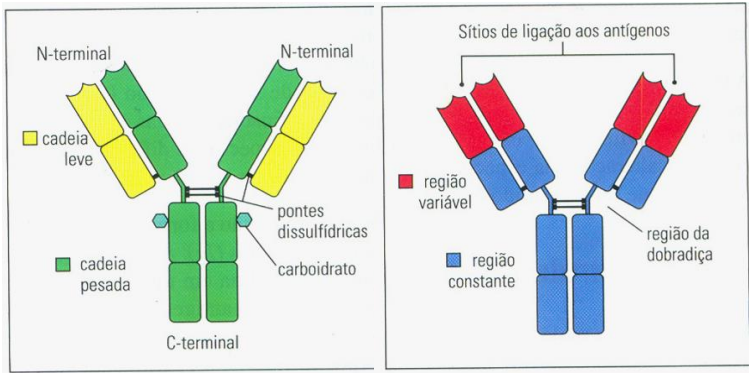
Os anticorpos IgG são constituídos de quatro cadeias polipeptídicas - duas cadeias pesadas idênticas e duas cadeias leves idênticas. Cada cadeia possui uma região V que contribui para o sítio de ligação do antígeno e uma região C que, na cadeia pesada, determina o isotipo do anticorpo e suas funções efetoras especializadas. A molécula de IgG possui a forma de Y, onde a haste e os braços têm tamanhos comparáveis e são flexíveis.

A digestão com a protease vegetal papaína produz três fragmentos, correspondentes aos dois braços e a haste. Os fragmentos correspondentes aos braços são chamados de Fab (de Fragment antigen binding) porque ligam-se ao antígeno. O fragmento correspondente a haste é chamado de Fc (de Fragmento cristalizável) porque, no primeiro experimento desse tipo, ele cristalizou. Por isso, a haste de uma molécula de anticorpo completa é conhecida como Região Fc ou Porção Fc e os braços como Fab. A dobradiça da molécula de IgG permite que os dois Fabs adotem muitas posições relativas diferentes entre si. Essa flexibilidade permite que antígenos distribuídos em distâncias distintas na superfície dos patógenos sejam ligados pelos dois braços Fab de uma molécula de IgG.

Diferenças nas regiões C da cadeia pesada definem cinco isotipos ou classes de imunoglobulinas, que possuem funções diferentes na resposta imune. São eles a imunoglobulina G (lgG), a imunoglobulina M (lgM), a imunoglobulina D (lgD), a imunoglobulina A (lgA) e a imunoglobulina E (lgE). Suas cadeias pesadas são representadas pela letra grega minúscula correspondente (γ, µ, δ, α e ε, respectivamente). A cadeia leve possui somente dois isotipos, denominados kappa (κ) e lambda (λ). Nenhuma diferença funcional foi encontrada entre anticorpos que possuem cadeias leves κ e os que portavam cadeias leves λ. Cadeias leves dos dois isotipos são encontradas associadas com qualquer isotipo de cadeias pesadas. Cada anticorpo, porém, contém cadeias leves κ ou cadeias leves λ, mas nunca ambas.

A estrutura de um único domínio de imunoglobulina pode ser comparada a um sanduíche, no qual duas folhas β (as fatias de pão) são mantidas juntas por fortes interações hidrofóbicas entre as cadeias laterais dos aminoácidos constituintes (o recheio). A estrutura é estabilizada por uma ponte dissulfeto entre as duas folhas β. As fitas adjacentes, nas folhas β são conectadas por alças da cadeia polipeptídica. Esse arranjo das folhas β fornece um arcabouço estrutural estável em todos os domínios de imunoglobulinas, já a sequência de aminoácidos das alças pode variar para conferir diferentes propriedades de ligação ao domínio.

A região V, na extremidade amino-terminal de cada cadeia pesada ou leve, é composta de um único domínio variável (domínio V), Vʜ na cadeia pesada e Vʟ na cadeia leve. Juntos, um domínio Vʜ e um domínio Vʟ formam o sítio de ligação do antígeno. Os outros domínios possuem pouca ou nenhuma diversidade de sequência em uma mesma classe de anticorpo e são denominados domínios constantes (domínios C), que compõem as regiões C. A região constante de uma cadeia leve é constituída de um único domínio Cʟ, ao passo que a região constante de uma cadeia pesada é composta de três ou quatro domínios C, dependendo do isotipo. As cadeias γ da IgG possuem três domínios, Cʜ1, Cʜ2 e Cʜ3. Alguns outros isotipos possuem quatro domínios C. Na molécula de IgG completa, o pareamento das quatro cadeias polipeptídicas produz três regiões globulares, correspondendo aos dois braços Fab e a haste Fc, sendo cada uma delas composta por quatro domínios de imunoglobulinas.

Cada braço contém um sítio de ligação do antígeno, e a haste contém os sítios de ligação para as células e moléculas efetoras, que permitem a eliminação do antígeno ligado do organismo. As cadeias de imunoglobulinas são constituídas por uma série de domínios de imunoglobulinas estruturalmente relacionadas. Nos domínios V, a variabilidade de sequências está localizada em três regiões hipervariáveis, correspondendo a três alças que estão agrupadas em uma extremidade do domínio. Na molécula do anticorpo, as alças hipervariáveis da cadeia pesada e leve formam uma superfície variável que liga o antígeno. O tipo de antígeno ligado por um anticorpo depende da forma do sítio de ligação do antígeno: antígenos que são moléculas pequenas podem se ligar em bolsas profundas; epítopos lineares de proteínas ou carboidratos podem ser ligados em fendas ou cristas; e a ligação de epítopos conformacionais de proteínas dobradas ocorre ao longo de toda a superfície. Anticorpos monoclonais são anticorpos de especificidade única, que são derivados de um clone de células produtoras de anticorpos idênticos, os quais são usados em testes diagnósticos e como agentes terapêuticos.



No genoma humano, os genes das cadeias pesadas e leves das imunoglobulinas estão em uma forma que não pode ser expressa. Nas células B em desenvolvimento, porém, esses genes sofrem rearranjos estruturais que permitem sua expressão. Os domínios V de cadeia leve e de cadeia pesada de imunoglobulina são codificados por dois (V e J) ou três (V. D e J) segmentos gênicos diferentes que são justapostos por reações de recombinação. Um mecanismo que contribui para a diversidade nas sequências na região V é a combinação aleatória de diferentes segmentos V e J nos genes de cadeia leve e de diferentes segmentos V, D e J nos genes de cadeia pesada, rearranjados. Um segundo mecanismo é a introdução de nucleotídeos adicionais (nucleotídeos P e N) nas junções entre os segmentos gênicos, durante o processo de recombinação. Um terceiro mecanismo que cria diversidade no sítio de ligação do antígeno, nos anticorpos, é a associação das cadeias leves e pesadas em diferentes combinações. O rearranjo gênico em uma célula B individual é estritamente controlado, de modo que apenas um tipo de cadeia pesada e um tipo de cadeia leve sejam expressos, resultando em uma célula B exclusiva, e sua progênie irá expressar imunoglobulina com uma única especificidade antigênica. Uma célula B virgem, que nunca encontrou antígeno, expressa somente imunoglobulina ligada à membrana das classes IgM e IgD. As cadeias pesadas µ e δ são produzidas a partir de uma única unidade transcricional, que sofre processamentos e reuniões alternativos de mRNA.










As imunoglobulinas ligadas a membranas, das classes IgM e IgD, atuam como receptores de antígenos ou receptores de células B, para células B virgens, maduras. Quando um patógeno se liga a um receptor de célula B, ela recebe sinais que causam proliferação e diferenciação celular de algumas das células B resultantes em células plasmáticas. Embora não possuam imunoglobulina em suas superfícies, as células plasmáticas secretam grandes quantidades de anticorpo IgM solúvel e algum IgD. A troca de imunoglobulina associada à membrana para anticorpo secretado é causada por um processamento alterado do mRNA de cadeia pesada, que elimina a sequência que codifica a região hidrofóbica de ancoramento à membrana. Outros membros do clone de células B em proliferação sofrem hipermutação somática, que introduz mutações ao longo das regiões V das cadeias leves e pesadas. Após a hipermutação, as células B portadoras de imunoglobulinas de superfície com afinidade mais alta pelos antígenos são selecionadas para se tornarem células plasmáticas. À medida que aumenta a afinidade pelo anticorpo, a dependência de IgM pentamérica por uma ligação mais forte com o antígeno é reduzida. Então se torna vantajoso mudar a imunoglobulina de IgM/lgD para um outro isotipo, IgG, IgE ou lgA, que tem funções efetoras mais adequadas ao tipo de patógeno infectante e ao sítio anatômico da infecção. Esse processo de troca de isotipo ocorre por meio de novas recombinações somáticas do gene de cadeia pesada expresso, onde o segmento VDJ rearranjado é movido para próximo de uma "nova" região C, com a excisão dos genes C µ e δ . Tanto a hipermutação somática quanto a troca de isotipo dependem da citidina desaminase induzida por ativação, uma enzima altamente específica de imunidade adaptativa que é produzida somente pelas células B que estão em proliferação em resposta a um antígeno.







Resumo
1 - A base estrutural da diversidade dos anticorpos
A - estrutura;
B - domínios proteicos;
C- sítio de ligação ao antígeno;
• constituição propriedades físicas.
D - produção de anticorpos.
2 - A geração da diversidade de Imunoglobulinas nas células B antes do encontro com o antígeno
1 - recombinação;
2 - ação das enzimas de recombinação;
3 - associação de cadeias leves e pesadas.
3 - Diversificação dos anticorpos após o encontro das células B com o antígeno
1 - hipermutação somática;
2 - troca de isotipo;
3 - diversidade das funções efetoras.
Referências
Parham, Peter et al. O sistema imune. 3. ed.-. Porto Alegre: ARTMED, 2011.



































Nenhum comentário:
Postar um comentário